Peptider er en klasse av forbindelser dannet ved kobling av flere aminosyrer gjennom peptidbindinger.De er allestedsnærværende i levende organismer.Til nå har titusenvis av peptider blitt funnet i levende organismer.Peptider spiller en viktig rolle i å regulere de funksjonelle aktivitetene til ulike systemer, organer, vev og celler og i livsaktiviteter, og brukes ofte i funksjonell analyse, antistoffforskning, medikamentutvikling og andre felt.Med utviklingen av bioteknologi og peptidsynteseteknologi har flere og flere peptidmedisiner blitt utviklet og brukt i klinikken.
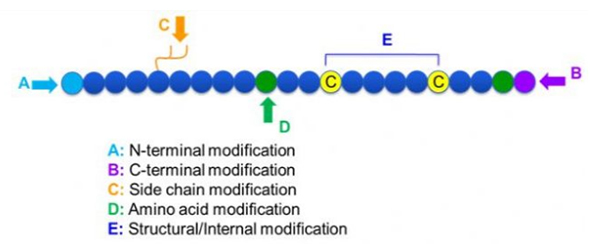
Det finnes et bredt utvalg av peptidmodifikasjoner, som enkelt kan deles inn i postmodifikasjon og prosessmodifikasjon (ved bruk av avledet aminosyremodifikasjon), og N-terminal modifikasjon, C-terminal modifikasjon, sidekjedemodifikasjon, aminosyremodifikasjon, skjelettmodifikasjon, etc., avhengig av modifikasjonsstedet (figur 1).Som et viktig middel for å endre hovedkjedestrukturen eller sidekjedegruppene til peptidkjeder, kan peptidmodifisering effektivt endre de fysiske og kjemiske egenskapene til peptidforbindelser, øke vannløseligheten, forlenge virkningstiden in vivo, endre deres biologiske fordeling, eliminere immunogenisitet , redusere toksiske bivirkninger, etc. I denne artikkelen introduseres flere viktige peptidmodifikasjonsstrategier og deres egenskaper.
1. Sykling
Sykliske peptider har mange anvendelser innen biomedisin, og mange naturlige peptider med biologisk aktivitet er sykliske peptider.Fordi sykliske peptider har en tendens til å være mer stive enn lineære peptider, er de ekstremt motstandsdyktige mot fordøyelsessystemet, kan overleve i fordøyelseskanalen og viser en sterkere affinitet for målreseptorer.Cyclization er den mest direkte måten å syntetisere sykliske peptider på, spesielt for peptider med stort strukturelt skjelett.I henhold til cykliseringsmodusen kan den deles inn i sidekjede-sidekjedetype, terminal - sidekjedetype, terminal - terminaltype (ende-til-ende-type).
(1) sidekjede-til-sidekjede
Den vanligste typen sidekjede til sidekjede-syklisering er disulfidbro mellom cysteinrester.Denne cykliseringen introduseres ved at et par cysteinrester avbeskyttes og deretter oksideres for å danne disulfidbindinger.Polysyklisk syntese kan oppnås ved selektiv fjerning av sulfhydrylbeskyttelsesgrupper.Cyclisering kan gjøres enten i et post-dissosiasjonsløsningsmiddel eller på en pre-dissosiasjonsharpiks.Cyclisering på harpikser kan være mindre effektiv enn løsemiddelsyklisering fordi peptidene på harpikser ikke lett danner cyklifiserte konformasjoner.En annen type sidekjede-sidekjedesyklisering er dannelsen av en amidstruktur mellom en asparaginsyre- eller glutaminsyrerest og baseaminosyren, som krever at sidekjedebeskyttelsesgruppen må kunne fjernes selektivt fra polypeptidet enten på harpiksen eller etter dissosiasjon.Den tredje typen sidekjede-sidekjede-cyklisering er dannelsen av difenyletere av tyrosin eller p-hydroksyfenylglycin.Denne typen cyklisering i naturprodukter finnes kun i mikrobielle produkter, og cykliseringsprodukter har ofte potensiell medisinsk verdi.Fremstillingen av disse forbindelsene krever unike reaksjonsbetingelser, så de brukes ikke ofte i syntesen av konvensjonelle peptider.
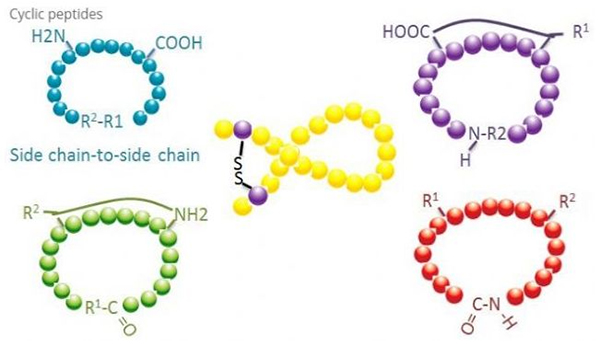
(2) terminal-til-sidekjede
Terminalsidekjedesyklisering involverer vanligvis C-terminalen med aminogruppen til lysin- eller ornitinsidekjeden, eller N-terminalen med asparaginsyre- eller glutaminsyresidekjeden.Annen polypeptidsyklisering gjøres ved å danne eterbindinger mellom terminal C og serin- eller treoninsidekjeder.
(3) Terminal eller head-to-tail type
Kjedepolypeptider kan enten sykles i et løsningsmiddel eller fikseres på en harpiks ved sidekjedesykling.Lave konsentrasjoner av peptider bør brukes i løsningsmiddelsentralisering for å unngå oligomerisering av peptider.Utbyttet av et hode-til-hale syntetisk ringpolypeptid avhenger av sekvensen til kjedepolypeptidet.Derfor, før du forbereder sykliske peptider i stor skala, bør det først opprettes et bibliotek med mulige kjedede blypeptider, etterfulgt av cyklisering for å finne sekvensen med de beste resultatene.
2. N-metylering
N-metylering forekommer opprinnelig i naturlige peptider og introduseres i peptidsyntese for å forhindre dannelse av hydrogenbindinger, og dermed gjøre peptider mer motstandsdyktige mot biologisk nedbrytning og clearance.Syntese av peptider ved bruk av N-metylerte aminosyrederivater er den viktigste metoden.I tillegg kan Mitsunobu-reaksjon av N-(2-nitrobenzensulfonylklorid) polypeptid-harpiks-mellomprodukter med metanol også brukes.Denne metoden har blitt brukt til å fremstille sykliske peptidbiblioteker som inneholder N-metylerte aminosyrer.
3. Fosforylering
Fosforylering er en av de vanligste post-translasjonelle modifikasjonene i naturen.I menneskelige celler er mer enn 30 % av proteinene fosforylert.Fosforylering, spesielt reversibel fosforylering, spiller en viktig rolle i å kontrollere mange cellulære prosesser, som signaltransduksjon, genuttrykk, cellesyklus og cytoskjelettregulering, og apoptose.
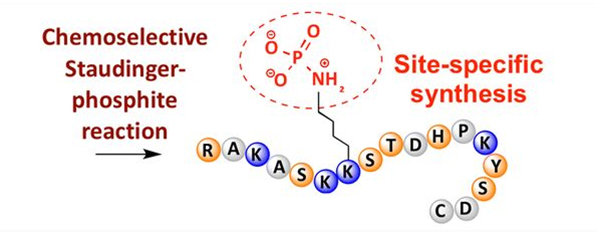
Fosforylering kan observeres ved en rekke aminosyrerester, men de vanligste fosforyleringsmålene er serin-, treonin- og tyrosinrester.Fosfotyrosin-, fosfotreonin- og fosfoserinderivater kan enten introduseres i peptider under syntese eller dannes etter peptidsyntese.Selektiv fosforylering kan oppnås ved å bruke rester av serin, treonin og tyrosin som selektivt fjerner beskyttende grupper.Noen fosforyleringsreagenser kan også introdusere fosforsyregrupper i polypeptidet ved postmodifikasjon.De siste årene har stedsspesifikk fosforylering av lysin blitt oppnådd ved bruk av en kjemisk selektiv Staudinger-fosfittreaksjon (Figur 3).
4. Myristoylering og palmitoylering
Acylering av N-terminalen med fettsyrer gjør at peptider eller proteiner binder seg til cellemembraner.Den myridamoylerte sekvensen på N-terminalen gjør det mulig å målrette Src-familiens proteinkinaser og revers transkriptase Gaq-proteiner for å binde seg til cellemembraner.Myristinsyre ble koblet til N-terminalen av harpiks-polypeptidet ved bruk av standard koblingsreaksjoner, og det resulterende lipopeptidet kunne dissosieres under standardbetingelser og renses ved RP-HPLC.
5. Glykosylering
Glykopeptider som vankomycin og teicolanin er viktige antibiotika for behandling av medikamentresistente bakterieinfeksjoner, og andre glykopeptider brukes ofte for å stimulere immunsystemet.I tillegg, siden mange mikrobielle antigener er glykosylerte, er det av stor betydning å studere glykopeptider for å forbedre den terapeutiske effekten av infeksjon.På den annen side har det blitt funnet at proteinene på cellemembranen til tumorceller viser unormal glykosylering, noe som gjør at glykopeptider spiller en viktig rolle i kreft- og tumorimmunforsvarsforskning.Glykopeptider fremstilles ved Fmoc/t-Bu-metoden.Glykosylerte rester, som treonin og serin, blir ofte introdusert i polypeptider av pentafluorfenolesteraktiverte fMOC-er for å beskytte glykosylerte aminosyrer.
6. Isopren
Isopentadienylering skjer på cysteinrester i sidekjeden nær C-terminalen.Proteinisopren kan forbedre cellemembranaffiniteten og danne protein-protein-interaksjon.Isopentadienerte proteiner inkluderer tyrosinfosfatase, liten GTase, cochaperone-molekyler, nukleær lamina og sentromere bindende proteiner.Isoprenpolypeptider kan fremstilles ved å bruke isopren på harpikser eller ved å introdusere cysteinderivater.
7. Modifikasjon av polyetylenglykol (PEG).
PEG-modifikasjon kan brukes til å forbedre proteinhydrolytisk stabilitet, biodistribusjon og peptidløselighet.Innføringen av PEG-kjeder til peptider kan forbedre deres farmakologiske egenskaper og også hemme hydrolysen av peptider av proteolytiske enzymer.PEG-peptider passerer lettere gjennom det glomerulære kapillærtverrsnittet enn vanlige peptider, noe som reduserer renal clearance.På grunn av den forlengede aktive halveringstiden til PEG-peptider in vivo, kan det normale behandlingsnivået opprettholdes med lavere doser og mindre hyppige peptidmedisiner.Imidlertid har PEG-modifikasjon også negative effekter.Store mengder PEG hindrer enzymet i å bryte ned peptidet og reduserer også peptidets binding til målreseptoren.Men PEG-peptiders lave affinitet oppveies vanligvis av deres lengre farmakokinetiske halveringstid, og ved å være tilstede i kroppen lenger, har PEG-peptider større sannsynlighet for å bli absorbert i målvev.Derfor bør PEG-polymerspesifikasjoner optimaliseres for optimale resultater.På den annen side akkumuleres PEG-peptider i leveren på grunn av redusert renal clearance, noe som resulterer i makromolekylært syndrom.Derfor må PEG-modifikasjoner utformes mer nøye når peptider brukes til medikamenttesting.
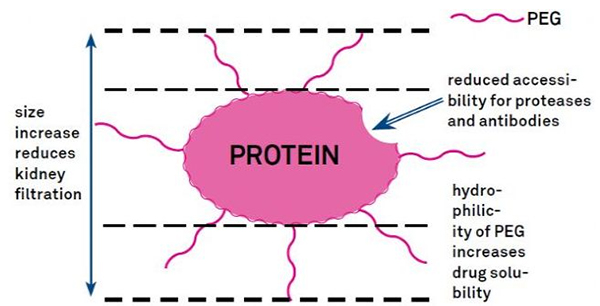
Vanlige modifikasjonsgrupper av PEG-modifikatorer kan grovt oppsummeres som følger: Amino (-amin) -NH2, aminometyl-Ch2-NH2, hydroksy-OH, karboksy-Cooh, sulfhydryl (-Tiol) -SH, Maleimid -MAL, succinimidkarbonat - SC, succinimidacetat -SCM, succinimidpropionat -SPA, n-hydroksysuccinimid -NHS, akrylat-ch2ch2cooh, aldehyd -CHO (som propional-ald, butyrALD), akrylbase (-akrylat-acrl), azido-azid, biotinyl Biotin, Fluorescein, glutaryl-GA, Acrylate Hydrazide, alkyn-alkyn, p-toluensulfonate -OTs, succinimidsuccinat -SS, etc. PEG-derivater med karboksylsyrer kan kobles til n-terminale aminer eller lysinsidekjeder.Aminoaktivert PEG kan kobles til asparaginsyre eller glutaminsyresidekjeder.Mal-aktivert PEG kan konjugeres til merkaptan av fullstendig avbeskyttede cysteinsidekjeder [11].PEG-modifikatorer er vanligvis klassifisert som følger (merk: mPEG er metoksy-PEG, CH3O-(CH2CH2O)n-CH2CH2-OH):
(1) rettkjedet PEG-modifikator
mPEG-SC, mPEG-SCM, mPEG-SPA, mPEG-OTs, mPEG-SH, mPEG-ALD, mPEG-butyrALD, mPEG-SS
(2) bifunksjonell PEG-modifikator
HCOO-PEG-COOH, NH2-PEG-NH2, OH-PEG-COOH, OH-PEG-NH2, HCl·NH2-PEG-COOH, MAL-PEG-NHS
(3) forgrening PEG-modifikator
(mPEG)2-NHS, (mPEG)2-ALD, (mPEG)2-NH2, (mPEG)2-MAL
8. Biotinisering
Biotin kan være sterkt bundet med avidin eller streptavidin, og bindingsstyrken er til og med nær kovalent binding.Biotinmerkede peptider brukes ofte i immunoassay, histocytokjemi og fluorescensbasert flowcytometri.Merkede antibiotinantistoffer kan også brukes til å binde biotinylerte peptider.Biotinmerker er ofte festet til lysinsidekjeden eller N-terminalen.6-aminokapronsyre brukes ofte som en binding mellom peptider og biotin.Bindingen er fleksibel i binding til underlaget og binder seg bedre i nærvær av sterisk hindring.
9. Fluorescerende merking
Fluorescerende merking kan brukes til å spore polypeptider i levende celler og for å studere enzymer og virkningsmekanismer.Tryptofan (Trp) er fluorescerende, så det kan brukes til egenmerking.Emisjonsspekteret til tryptofan avhenger av det perifere miljøet og avtar med synkende løsningsmiddelpolaritet, en egenskap som er nyttig for å påvise peptidstruktur og reseptorbinding.Tryptofanfluorescens kan slukkes av protonert asparaginsyre og glutaminsyre, noe som kan begrense bruken.Dansylkloridgruppen (Dansyl) er svært fluorescerende når den er bundet til en aminogruppe og brukes ofte som en fluorescerende etikett for aminosyrer eller proteiner.
Fluorescensresonans Energikonvertering (FRET) er nyttig for enzymstudier.Når FRET påføres, inneholder substratpolypeptidet vanligvis en fluorescensmerkende gruppe og en fluorescensdempende gruppe.Merkede fluorescerende grupper slukkes av quencheren gjennom ikke-fotonenergioverføring.Når peptidet dissosieres fra det aktuelle enzymet, avgir merkegruppen fluorescens.
10. Burpolypeptider
Burpeptider har optisk fjernbare beskyttende grupper som beskytter peptidet fra å binde seg til reseptoren.Når det utsettes for UV-stråling, aktiveres peptidet, og gjenoppretter dets affinitet til reseptoren.Fordi denne optiske aktiveringen kan kontrolleres i henhold til tid, amplitude eller plassering, kan burpeptider brukes til å studere reaksjoner som oppstår i celler.De mest brukte beskyttelsesgruppene for burpolypeptider er 2-nitrobenzylgrupper og deres derivater, som kan introduseres i peptidsyntese via beskyttende aminosyrederivater.Aminosyrederivater som er utviklet er lysin, cystein, serin og tyrosin.Aspartat- og glutamatderivater brukes imidlertid ikke ofte på grunn av deres mottakelighet for cyklisering under peptidsyntese og dissosiasjon.
11. Polyantigenisk peptid (MAP)
Korte peptider er vanligvis ikke immune og må kobles til bærerproteiner for å produsere antistoffer.Polyantigenisk peptid (MAP) er sammensatt av flere identiske peptider koblet til lysinkjerner, som spesifikt kan uttrykke immunogener med høy styrke og kan brukes til å fremstille peptid-bærerproteinkupletter.MAP-polypeptider kan syntetiseres ved fastfasesyntese på MAP-harpiks.Imidlertid resulterer ufullstendig kobling i manglende eller avkortede peptidkjeder på noen grener og viser derfor ikke egenskapene til det originale MAP-polypeptidet.Som et alternativ kan peptider fremstilles og renses separat og deretter kobles til MAP.Peptidsekvensen festet til peptidkjernen er veldefinert og lett karakterisert ved massespektrometri.
Konklusjon
Peptidmodifikasjon er et viktig middel for å designe peptider.Kjemisk modifiserte peptider kan ikke bare opprettholde høy biologisk aktivitet, men også effektivt unngå ulempene med immunogenisitet og toksisitet.Samtidig kan kjemisk modifikasjon gi peptider noen nye utmerkede egenskaper.I de senere årene har metoden for CH-aktivering for post-modifisering av polypeptider blitt raskt utviklet, og mange viktige resultater har blitt oppnådd.
Innleggstid: 20. mars 2023